

Рождение ДНК-генеалогии в 2008 году и последующее развитие событий с неизбежностью показывало, что коллизия с популяционной генетикой, или попгенетикой, как я буду далее называть для краткости, неизбежна. На самом деле не с «попгенетикой», конечно, а с ее руководителями и исполнителями. Популяционная генетика человека уже давно, собственно, с 1990-х годов, фактически превратилась в псевдонауку, особенно, в отношении ее расчетного аппарата и соответствующих экскурсов в историю в виде «интерпретаций».
У попгенетиков со временем развилось ощущение безнаказанности в отношении того, что чтобы бы они ни написали, какую бы цифру ни вбросили в академические публикации, какую «теорию» как якобы проверенную и подтвержденную ни опубликовали, никто не будет ни проверять, ни подвергать сомнению, все это пойдет в печать. Ситуация усугубилась после 2004 года, когда в попгенетике появился так называемый «метод Животовского», оперирующий некими «популяционными скоростями», в корне неверными, и приводящими к завышениям датировок древних событий на 250-350% и выше. Метод был провозглашен как «откалиброванный» по древним генеалогиям, но первое же рассмотрение показало, что за «калибровку» выдавалась откровенная подгонка, манипуляция данными, подтасовки цифр. За прошедшие с тех пор десять лет академические журналы в области попгенетики человека оказались наводнены фактически мусором, не выдерживающим никакой критики.
Начиная с 2008 года, ДНК-генеалогия, базирующаяся на принципах физической химии и химической кинетики, то есть науки о скоростях и механизмах химических и биологических реакций, систематически занялась выявлением наиболее значимых ошибок, подтасовок, манипуляций, искаженных датировок и соответствующих исторических «интерпретаций» попгенетиками. Этому были посвящены многочисленные публикации в научных журналах, неподконтрольных попгенетикам. Попытки публиковать дискуссионные статьи в «подконтрольных» журналах немедленно пресекались без каких-либо попыток научного разбора, мотиваций, объяснений. В недавней книге «Происхождение славян» (А.А. Клёсов, Москва, 2013) целая глава объемом более 100 страниц под названием «Берегитесь популяционных генетиков в исторических «исследованиях»» была посвящена этой теме, с многочисленными примерами. Ни одного комментария в научной печати, ни одной попытки ответить на вызов ДНК-генеалогии сделано не было с 2009 года, когда на мой разгромный комментарий на 14 страницах в одном из ведущих журналов мира Human Genetics в отношении ведущих попгенетиков мира и их дефектной методологии они ответили вялым, пассивным комментарием. Он сводился к тому, что их «откалиброванная» методология опубликована и активно используется в популяционной генетике, «пройдя проверку», и что подход ДНК-генеалогии «очень интересен», но к нему «следует относиться с осторожностью». С тех пор, за прошедшие пять лет, никаких других рассмотрений критики ДНК-генеалогии не было. Видимо, попгенетики полагали, что «само рассосется». Не рассосалось.
Игнорировать критику дальше попгенетики не могли, разобраться в своих принципиальных ошибках не смогли или не захотели, на контакты не шли, более того, контакты отвергали. Для них оставалось только ответить агрессией, огульными обвинениями, инсинуациями, без каких-либо попыток разобраться в вопросе. Эта агрессия свершилась в конце ноября 2014 года на конференции «Этногенез, история, язык и культура карачаево-балкарского народа», проводимой в РАН, на Ленинском проспекте в Москве. Это была скоординированная акция, для которой все средства были хороши. В конференции принимали участие два попгенетика из РАН, и два – из РАМН, и все четверо вместо докладов по тематике Конференции сделали доклады персональной направленности, полностью направленные на дискредитацию нового научного направления и меня лично – с показом портретов (Боринская, Балановская, Балановский) и выдачей персональных негативных комментариев, заполнявших многочисленные слайды. Ни один из них не касался собственно науки, вместо этого передергивали цитаты из моих научных и популярных докладов, не имеющих отношения к ДНК-генеалогии и вообще ни к какой науке, вытаскивали (смехотворный) «компромат» на издательство, опубликовавшее мою книгу, приводили не мои цитаты, выдавая их за якобы мои.
В итоге, после попытки откровенной обструкции и срыва моей лекции, на которую собрались участники конференции, мать и сына Балановских удалили со сцены и изгнали из зала заседаний под одобрительные возгласы аудитории. Больше они на конференции не появлялись. Но в тот же вечер в СМИ появились лживые статьи, под названиями «Лжегенетик нахимичил» и «Провал ДНК-генеалогии в РАН». Балановской в тот же день была составлена фальшивая «Резолюция Круглого стола», выставлена на сайте ее лаборатории в РАМН, размещена на «моей» странице в Википедии. Она принципиально отличалась от текста настоящей Резолюции, зачитанной на заключительном пленарном заседании Конференции. Только через 10 дней тот же подметный тест «резолюции» был направлен со стороны Балановской «для согласования» в оргкомитет и докладчикам. Естественно, я категорично возразил против лживых формулировок этой «резолюции». Ответа не последовало ни от Балановской, ни от Оргкомитета. Так что пока результат проталкивания Балановской фальшивого текста «резолюции» неизвестен.
Чтобы внести ясность в ситуацию и в суть коллизии ДНК-генеалогии и популяционной генетики, я написал настоящую статью. Статья будет размещена на сайте «Переформат» и после этого на других сайтах, будет направлена в Президиум РАН, в сжатом виде будет направлена для публикации в журнал «Биохимия» Российской АН, и переведена для публикации в зарубежных изданиях.
Предисловие
Шесть лет назад, в 2008 году, ДНК-генеалогия заявила о себе как о новой научной дисциплине. Начало было положено обширным анализом в рамках новой науки в то время протяженных (25-маркерных) гаплотипов гаплогруппы R1a Y-хромосомы европейского мужского населения, с расчетом времен жизни общих предков по странам и регионам (результаты были опубликованы в третьем выпуске журнала «Вестник Российской Академии ДНК-генеалогии» в августе 2008 года). Было показано, что мужское население (гаплогруппы R1a) современной Германии имеет общего предка примерно 4600 лет назад, и базовый (предковый) гаплотип его был следующий:
13 25 16 10 11 14 12 12 10 13 11 30 15 9 10 11 11 24 14 20 32 12 15 15 16
25 ноября того же года, то есть через три месяца, вышла статья Haak et al. (PNAS, 2008), в которой были описаны ископаемые гаплотипы гаплогруппы R1a, найденные в местечке Эулау, Германия, с датировкой 4600 лет назад. Ископаемые гаплотипы имели вид
13/14 25 16 11 11 14 X X 10 12/13 X 30 14/15 X X X X X 14 19 X X X X X
(двойные числа в ископаемых гаплотипах – это те, которые точно определить не смогли, здесь возможны варианты). Знаки X указывают на пропущенные аллели в ископаемых гаплотипах. Только две аллели (числа в гаплотипах выше) различались на один шаг, 10 и 11 в четвертом положении (DYS391), и 19 и 20 ближе к концу гаплотипа (DYS 448). Это вполне естественно, никто не мог утверждать, что найденные в захоронении останки принадлежали именно первопредку «германских» R1a, тем более что там была целая семья. Две мутации разницы по расчетам в рамках ДНК-генеалогии дают всего несколько сотен лет расстояния между ними, что укладывается в диапазон погрешности расчетов (Klyosov, J. Gen. Genealogy, 2009).
Со времени этого первого положительного испытания и демонстрации практического совпадения расчетов и археологических датировок, то есть с лета 2008 года, и начался отсчет новой науки ДНК-генеалогии.
Чтобы не быть голословным, покажу, как проводился расчет и на основании чего. В те времена, 2008-й год, в академической литературе использовались в основном гаплотипы от 6- до 10-маркерных, были известны 12-маркерный (FTDNA) и 17-маркерный гаплотип (последний был введен в 2006 году), но для хронологических расчетов они практически не использовались. Короткие же гаплотипы обрабатывались популяционными генетиками с применением «популяционной» скорости мутации, которую я опишу в подробностях позже. Здесь скажу лишь то, что она ужасающе неверна, и завышает датировки обыкновенно на 250-350%.
В подходе ДНК-генеалогии 2008 года строилось дерево гаплотипов для проверки на общую симметричность, то есть на отсутствие выраженных ветвей.

Рис. 1. Дерево из 67 25-маркерных гаплотипов
современных немцев гаплогруппы R1a (Клёсов, 2008).
Дерево выше показывает общую симметричность (множество мелких ветвей в нем можно анализировать по отдельности, но опыт показывает, что итоговый результат будет практически такой же), и его можно количественно обрабатывать как единую систему, то есть что все 67 гаплотипов происходят от одного общего предка. На все 67 гаплотипов в 12-маркерном формате имеется 208 мутаций от приведенного выше базового гаплотипа, и в 25-маркерном формате – 488 мутаций. В первом случае получаем 208/67/0.02 = 155 → 183 условных поколений (по 25 лет) от общего предка (стрелка указывает на поправку на возвратные мутации), то есть 4575±560 лет до общего предка. Во втором случае, для 25-маркерных гаплотипов, получаем 488/67/0.046 = 158 → 187 условных поколений, то есть 4675±510 летдо общего предка. Как видим, совпадение почти абсолютное для расчетов по 12- или 25-маркерным гаплотипам. Здесь (и вообще в ДНК-генеалогии) на поколение берется 25 лет, это – математическая величина, не имеющая отношения к продолжительности «бытового поколения», которое есть величина плавающая, и зависит от обычаев, религиозных верований, эпохи, мора, войн и так далее.
Условное поколение в 25 лет привязано к величине константы скорости мутации 0.02 (мутаций на условное поколение на 12-маркерный гаплотип), или 0.046 (мутаций на условное поколение на 25-маркерный гаплотип). Константы скорости мутации для более протяженных гаплотипов (37-, 67- и 111- маркерных) будут даны позже. Надо сказать, что уже в первом выпуске Вестника, в июне 2008 года, проводились многочисленные расчеты для 37- и 67-маркерных гаплотипов, что тогда было неслыханно. Результаты всех этих расчетов верны до настоящего времени.
Возможно, неопытный читатель тут же запротестует, мол, как можно по всей Германии, или любому другому региону судить по каким-то 67 гаплотипам, ведь там живут миллионы человек. На это я обычно задаю встречный вопрос – чтобы понять, что море соленое, нужно ли пробовать воду из всех бухточек? А как же химики делают анализ растворов, беря пробу всего лишь в долю миллилитра из ведра, цистерны, озера? Когда делают анализ крови, что, всю кровь из вас выкачивают? Нет. Значит, вопрос о размере пробы поставлен некорректно. Вопрос на самом деле сводится к тому, равномерно ли перемешана система, чтобы по малой пробе, или выборке, судить обо всей системе. Этот вопрос в ДНК-генеалогии давно проработан, и результаты сопоставительных исследований опубликованы (напр., Клёсов, Биохимия, 2011). В горах – одна ситуация, там в каждой долине состав гаплотипов (и гаплогрупп/субкладов) может заметно различаться, на равнине – другая, там гаплотипы более перемешаны за тысячелетия. Но в любом случае расчеты и выводы по гаплотипам относятся только к той конкретной выборке, которая рассматривалась, и только сопоставление разных выборок может показать, насколько система «перемешана», и решить вопрос об обобщении результатов расчетов и выводов на всю популяцию региона, или только ее конкретную часть.
Чтобы не быть голословным, рассмотрим выборку по Германии уже 2014 года. Теперь в ней 286 гаплотипов в 67- и 65 гаплотипов в 111-маркерном формате (база данных IRAKAZ-2014). Ниже – дерево 67-маркерных гаплотипов.
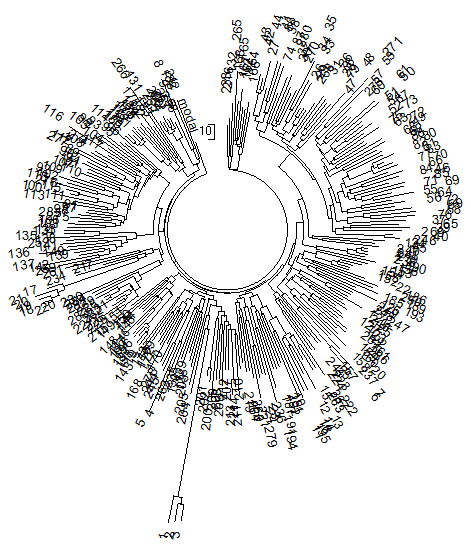
Рис. 2. Дерево из 286 67-маркерных гаплотипов современных немцев гаплогруппы R1a (база данных IRAKAZ-2014). Три гаплотипа внизу (номера 1, 2, 3) относятся к древнейшему субкладу гаплогруппы, R1a-M420, и в расчеты (см. текст) не включались. Расчеты проводили на калькуляторе Килина-Клёсова, поскольку ручной расчет системы с более чем 22 тысячи аллелей слишком трудозатратен.

Общий предок всех 67-маркерных гаплотипов жил 4848±415 лет назад, всех 67- и 111-маркерных гаплотипов – 4294±298 лет назад, при округлении получаем 4800±400 и 4300±300 лет назад. Среднюю величину можно не брать, и так видно, что она практически равна результатам, полученным ранее, в 2008 году, и с совершенно другой выборкой в Германии 4575±560 лет и 4675±510 лет до общего предка, расчитанным по 12- и 25-маркерным гаплотипам.
Таким образом, обе выборки – 2008 и 2014 гг. являются вполне репрезентативными для целей настоящего исследования.
Теперь посмотрим, как с этой задачей «справились» бы популяционные генетики. Вот так они считают в десятках, если не в сотнях статей, опубликованных в ведущих академических журналах мира. Они берут те же 208 мутаций (в 12-маркерных гаплотипах в даном случае), делят на число маркеров (они же локусы), которых во всех 67 12-маркерных гаплотипах 67х12 = 804 маркера, и далее делят на «популяционную скорость», равную 0.00069 мутаций на маркер на 25 лет. Заметим, что 25 лет здесь – это «просто так», типа поколение, никакого обоснования не дается. А если бы и давалось – ничего бы не изменило, снявши голову по волосам не плачут. Эта величина все равно неверна. Итак, вот как они считают:
208/804/0.00069 = 375.937 поколений, то есть 9373 лет до общего предка. Не 4600, как при расчетах ДНК-генеалогии, а 9373±3592 лет. То есть промах с датировкой примерно в два раза и огромная погрешность. Причем я не случайно здесь привел их датировку с точностью до третьего знака «после запятой», они так обычно записывают и публикуют. Их примитивное математическое образование не дает им сообразить, что если ошибка равна ±3592 (это уже смешно, ошибки с такой точностью не определяются), то есть уже в тысячах лет, то писать в главном числе 9373, то есть с точностью до года, не имеет никакого смысла. Это – безграмотно. Но они все так пишут. И никто им не скажет, что запись в лучшем случае должна быть в виде 9400±3600 лет, но и она неверна, должно быть 4600 лет с погрешностью, как определено методами ДНК-генеалогии, достоверность которых показана на множестве примеров (см. ниже).
Думаете, я шучу? Вовсе нет. Вот – самый недавний пример, статья попгенетиков «Tracing the genetic origin of Europe’s first farmers reveals insights into their social organization» (Szécsényi-Nagy, Brandt, Keerl et al., bioRxiv, 3 сентября 2014). Нашли двух «охотников-собирателей» (на самом деле «охотники-собиратели» – это у попгенетиков фигура речи, это просто те, кто жил ранее 7000 лет назад) среди 19 скелетных остатков. Показывают их долю, то есть «частоту» на языке попгенетиков: 2/19 = 10.53%. Любой человек с минимальным математическим образованием вздрогнет. Как можно для двух человек из 19 считать «частоту» с точностью до второго знака «после запятой»? Почему хотя бы не 11%, что тоже не очень правильно. Правильнее – 11±8%, и то при доверительном интервале всего лишь 68%. Потому что для двух единиц погрешность «частоты» там примерно ±71%. И вот имея 11±8%, они пишут 10.53%. Это и есть отсутствие минимальной научной школы у популяционных генетиков.
Подобные ошибки постоянно делают попгенетики Балановские. В статье про Кавказ (2011) они берут семь абхазов из 58 общей выборки, вычисляют «частоту» как 12.1%. А там должно быть 12±5%, какие десятые доли процента? В итоге цифры на самом деле получаются вовсе не такие точные, как авторы пытаются показать, что смазывает результаты работы. А дальше – пошло-поехало (таблица 5 статьи) – 1.44±0.63, 1.09±0.68, 1.77±0.99, 2.38±0.89, 2.45±0.76, и так далее. Это и есть безграмотность и отсутствие научной школы. Про расчеты времен до общих предков и говорить не приходится, об этом позже.
Возращаемся, брезгливо отряхиваясь, к ДНК-генеалогии. Помимо того, что в 2008 году ДНК-генеалогия прошла проверку на реальных (ископаемых) гаплотипах, показав ту же хронологию, в том же 2008 году, в июне, вышел первый выпуск Вестника, который позже стал международным. В первом выпуске были две больших статьи – «Основные правила ДНК-генеалогии» (54 страницы) и «Происхождение евреев с точки зрения ДНК-генеалогии» (179 страниц). С тех пор вышло 66 выпусков Вестника, общим объемом более 10 тысяч страниц, все выпуски в открытом доступе. Cо второго выпуска (июль 2008) в разделе «Обращения читателей и персональные случаи ДНК-генеалогии» публикуются ответы на письма читателей из разных стран мира. До сегодняшнего выпуска опубликованы персональные истории в виде ответов на 234 письма читателей. Авторы Вестника – в основном российские специалисты и любители ДНК-генеалогии, но также специалисты из США, Италии, Франции, Словении, Сербии, Италии, Германии, Бельгии, Шотландии и других стран.
С того же 2008 года опубликованы 17 научных статей по ДНК-генеалогии в академических журналах (все, кроме одной, на английском языке, и одна – в старейшем российском академическом журнале «Биохимия»), 6 книг по ДНК-генеалогии (5 на русском языке и одна на сербском, в Белграде), и 82 научно-популярных статьи по ДНК-генеалогии, из них 50 статей на Переформате – сайте, который выпускается профессиональными историками. Еще несколько статей на Переформате добавил член Академии ДНК-генеалогии И.Л. Рожанский, выпускник того же Химического факультета МГУ, что и автор настоящей статьи.
Академия ДНК-генеалогии была создана в том же 2008 году. Как известно, Академия – это сообщество научных единомышленников, выпускающих свое печатное издание. Пока Академия зарегистрирована только в США, но в настоящее время готовится к регистрации в России (Москва). Состав членов Академии не расширяется с 2008 года, и делается это намеренно, чтобы не «снижать планку» Академии. Два основных требования к членам Академии – знать свои гаплотип и гаплогруппу, и публиковаться в научных изданиях по теме ДНК-генеалогии. После регистрации в России состав членов Академии будет расширяться, и, видимо, значительно, вплоть до десятков и сотен человек.
Научные статьи в области ДНК-генеалогии публиковались в журналах Human Genetics (2009), J. Gen. Genealogy (2009), Advances in Anthropology (2011-2014), Биохимия (2011), Eur. J. Human Genetics (2014), Frontiers in Genetics (2014). О некоторых статьях ниже будет рассказано более подробно. Здесь отметим, что весной 2013 года я был приглашен быть главным редактором журнала Advances in Anthropology (Успехи антропологии), и сделал его профильным журналом по ДНК-генеалогии (наряду, конечно, с публикациями авторов по разным направлениям антропологии).
Первой статьей в АА была наша (совместно с И.Л. Рожанским) статья по константам скоростей мутаций в гаплотипах Y-хромосомы и их калибровке, то есть количественной связи с хронологией. Концептуально сделать это было несложно, поскольку кинетика химических и биологических мутаций – моя прямая специальность, я в свое время заканчивал кафедру академика Н.Н. Семенова, единственного лауреата Нобелевской премии среди советских химиков, и написал немало учебников для Высшей школы (в основном издательства Московского университета и издательства «Наука»), часть из них – с моим учителем, деканом Химического факультета МГУ чл.-корр. АН СССР И.В. Березиным. За разработку кинетики химических и биологических реакций я в 1984 году стал лауреатом Государственной премии СССР по науке и технике, а до того – лауреатом премии Ленинского комсомола (1978). Так что концептуально разработать количественную основу ДНК-генеалогии было несложно, но надо было ее верифицировать на тысячах примеров, чтобы она была надежной. Это было сделано с использованием 3160 гаплотипов (2489 из них – в 67-маркерном формате, из 55 ДНК-генеалогических линий из 8 основных гаплогрупп). Эта работа была опубликована в Advances in Anthropology в 2011 году, во втором выпуске журнала после его основания в том же году (Rozhanskii & Klyosov, v. 1, No. 2, 26-34, 2011). Именно за публикацию серии статей, значительно расширивших читательскую аудиторию, я и был приглашен занять пост главного редактора журнала, спустя два года после первых публикаций там.
В последующем в том же журнале были опубликованы статьи с детальным описанием гаплогруппы R1a в мире, гаплогруппы R1a в Европе (с описанием 38 ветвей гаплогруппы R1a), гаплогруппы R1b, ДНК-генеалогии и лингвистики, ДНК-генеалогии и древней керамике от трипольской культуры до культур Юго-Восточной Азии и обеих Америк, а также четыре статьи об Африке и происхождении человека с точки зрения ДНК-генеалогии (две в 2012 г, и две – в 2014 г, одна из них редакторская). Одна статья из этих четырех (Klyosov & Rozhanskii, 2012) установила своеобразный рекорд популярности у читателей журнала – по состоянию на 18 декабря 2014 года она была просмотрена 75891 раз, и «скачана» 16112 раз (ссылка).
Полагаю, что описанное выше – это крупный успех нового направления, отражение его признания. Поэтому более чем странно было увидеть резкий «накат» группы популяционных генетиков на ДНК-генеалогию на московской Конференции по карачаево-балкарцам, причем «накат» скоординированный, срежиссированный, заранее подготовленный, со сбором предупрежденных людей по спискам, и далее, когда акция провалилась, началась агрессивная и лживая кампания со стороны тех же популяционных генетиков, ведомых матерью и сыном Балановскими, с выставлением пасквилей в СМИ под заголовками «Провал ДНК-генеалогии в РАН», «Лжегенетик нахимичил» и подобных. С трибуны Конференции О. Балановским, зав. лабораторией Ин-та Общей генетики РАН было объявлено, что моя книга «Происхождение славян» (2013) была опубликована издательством («Алгоритм», Москва), которое также якобы публиковало труды Геббельса и Муссолини. Это, видимо, было призвано вызвать возмущение аудитории и намекнуть, что моя книга тоже фашистская. Это что, научная дискуссия? Где научная этика? За это раньше били канделябрами, и отнюдь не иносказательно.
Для справки, Издательство «Алгоритм» — российское книжное издательство, основано в 1996 году, специализируется на выпуске книг острой и актуальной общественно-политической и исторической направленности. Среди авторов опубликованных книг – Сергей Кара-Мурза, Александр Зиновьев, Олег Кашин, Сергей Глазьев, Дмитрий Рогозин, Михаил Делягин, Михаил Ходорковский, Анатолий Лукьянов, и, как сообщает издательство, многих других аналитиков, мыслителей, историков, философов, политологов, социологов, писателей… Издательство регулярно участвует в крупных российских и международных книжных выставках, является лауреатом премий и конкурсов в книгоиздательстве. Да, если издательство и публиковало книги Геббельса или Муссолини, для исторических целей, то при чем здесь другие авторы? Я, например, написал в 2000-х годах ряд аналитических работ о причинах и обстоятельствах прихода Гитлера к власти в 1932-1933 гг., и использовал для них опубликованные в советской печати дневники Геббельса, которые были полны отчаяния от ожидаемого развала коалиции, и готовность покончить жизнь самоубийством – и это в преддверии неожиданной победы Гитлера и его группы на выборах в Рейхстаг. Что здесь неправильного, когда историки используют эти работы?
Это только показывает ущербность О. Балановского и его страстное желание дискредитировать ДНК-генеалогию отнюдь не научными методами. Откуда такое страстное желание проф. О. Балановского, проф. Е. Балановской (лаборатория популяционной генетики РАМН), их коллег биолога С. Боринской (из того же Института общей генетики) и антропологов Д. Пежемского и С. Дробышевского (МГУ) дискредитировать, причем откровенно лживыми методами? Дробышевский, который, по его собственному признанию, в ДНК-генеалогии не разбирается и «генетику» не комментирует, объявил, что это «лженаука». Откуда это?
Об этом – настоящая статья, в нескольких частях.
Закончу предисловие тем, что уже в начале зарождения ДНК-генеалогии ее создатели обратили внимание на вопиющую ненаучность подходов популяционной генетики к расчету скоростей мутаций в Y-хромосоме ДНК. Никакого подразделения гаплогрупп на ветви не проводилось, расчеты велись «поперек» ветвей, каждая из которых имеет своего общего предка, и в итоге на свет появлялся некий «фантомный предок» со своей фантомной «хронологией», не имеющей ничего общего с реальностью. «Калибровка» этих подходов была возмутительной, фактически фальсификацией, при которой «мешающие» гаплотипы изымались, вводились некие подгоночные «коэффициенты», и так далее. Получаемый мусор, другого слова не подобрать, наводнял (и продолжает наводнять) академические журналы. Это была трагедия популяционной генетики. Было совершенно ясно, что у них нет никакой научной школы, при которой получаемая чепуха выявляется и останавливается на первом же научном семинаре.
Еще в 2008 году я попытался связаться с популяционными генетиками Л. Животовским и О. Балановским, но ответа не получил. Пришлось открыть критику в научной печати, с конкретными примерами и обоснованиями. Тем временем критика их «методов» нарастала на сетевых форумах, в блогах. Любители понимали в искажениях и ошибках попгенетиков несравненно больше, чем «специалисты». Но «специалистов» это не интересовало, у них были свои дела, куда разбор критики не входил. Они были заняты генерацией мусора в академические издания. Моя критика усилилась, в Вестнике появился раздел «Берегитесь популяционных генетиков» с многочисленными примерами их «мусора». Была подробно разобрана их «калибровка» скоростей мутаций, выявлена их полная ненаучность, с приведением множества примеров полной их некомпетентности в расчетах и исторических интерпретациях. У них был выход в том, чтобы связаться со мной и разобраться, что они делают не так. Очевидно, что этот научный путь им не подошел. Они решили «пойти свиньей».
На этом пути они буквально оголтело пошли во все тяжкие. Публичная ложь, подделка Резолюции Круглого стола (формулировки которой еще продолжали обсуждаться среди членов Оргкомитета и докладчиков 7 декабря, как описано в последующих частях этой статьи, иначе говоря, Резолюция еще не приобрела окончательный вид, но уже 27 ноября на вебсайте лаборатории Е. Балановской появилась якобы утвержденная и готовая резолюция с лживыми негативными словами про то, что «в рамках распространяемой в интернете «ДНК-генеалогии» отождествляются принципиально разные этнические и генетические категории, прикрепляются этнические названия к генетическим «родам» и археологическим культурам. Понятийно-расчётный аппарат ДНК-генеалогии научно необоснован, не прошёл научную апробацию в индексируемых научных журналах и основан на смешении популяционно-генетических, генеалогических и историко-культурных понятий. Все эти черты ДНК-генеалогии выводят ее за рамки академических наук и находятся в противоречии с необходимостью корректного подхода, различающего биологические данные о популяции от понятия народа во всей его сложности. Поэтому упоминание ДНК-генеалогии в названии Круглого стола неуместно».
Заметьте – «в рамках распространяемой в интернете «ДНК-генеалогии», то есть умышленно делается вид, что публикуемых книг и статей не существует, что не существует Круглого стола на Конференции, в названии которого присутствуют слова «ДНК-генеалогия». Далее, что «понятийно-расчетный аппарат ДНК-генеалогии научно необоснован» – что это, как не нахрапистая ложь? «Не прошел научную апробацию», «выводят за рамки академических наук», «находятся в противоречии с необходимостью корректного подхода», и так далее. То, что авторы этой «резолюции» были сильно нетрезвы, это вряд ли. Ими вела ненависть, сжигающая ненависть. И это – Институт общей генетики, который так пострадал во времена Т. Лысенко, а теперь, оказывается, история повторяется. Но уже в виде фарса, как известно.
Это – ненависть и отчаяние крысы, загнанной в угол. Выйти из угла у Балановских и иже с ними уже не получится, теперь я уже не дам, в виде научных и популярных статей в России и за рубежом, в виде книг. Или они покинут Академию, или публично покаются и начнут осваивать ДНК-генеалогию. Другого выхода, думаю, у них нет. А если и окажется, как вполне возможно в современной России, все равно будут жить в страхе текущих и последующих разоблачений.
Но всё, как известно, делается к лучшему. Гнойник у Балановских лопнул. И их истеричный «накат» теперь позволит мне последовательно объяснить – не в первый раз, но с учетом «наката» – принципы ДНК-генеалогии, независимость ее от попгенетики, сопоставить понятийные и расчетные аппараты, привести примеры подтасовок, махинаций, некомпетентности попгенетики в исполнении Балановских, и не только их, привести примеры того, как надо рассчитывать хронологию по снип-мутациям и мутациям в гаплотипах, и как складываются интерпретации корректно полученных данных – в рамках ДНК-генеалогии. И, напротив, какой получается мусор в исполнении популяционной генетики.
Приступим.
Популяционная генетика и ДНК-генеалогия как разные области наук:
методология ДНК-генеалогии – физическая химия и исторический анализ,
результат – «молекулярная история»
Основы ДНК-генеалогии уже созданы, причем стремительно (Klyosov, 2008а, 2008b, 2009c, 2009a, 2009b, 2009c, 2010; Клёсов, 2011; Rozhanskii, Klyosov, 2011, 2012; Klyosov, 2012; Klyosov, Rozhanskii, 2012a, 2012b; Klyosov et al., 2012; Klyosov, Tomezzoli, 2013; Клёсов, 2013а, 2013b; Klyosov, Mironova, 2013; Klyosov, 2014a, 2014b, 2014c; Tofanelli, Klyosov et al, 2014), и идет прогрессивное накопление экспериментального материала. Новый материал поступает потоком, ежедневно в базы данных уходят десятки и сотни новых «экспериментальных точек», которые по принципу обратной связи корректируют методологию новой науки, что приводит к уточнению методов расчета.
Экспериментальные данные ДНК-генеалогии — это картина мутаций в нерекомбинантных (то есть не рекомбинирующихся с другими хромосомами в ходе передачи наследственной информации потомкам) участках мужской половой хромосомы (на самом деле — и в митохондриальной ДНК, но в этой статье речь пойдет о Y-хромосоме, более информативной для исторических исследований), причем картина мутаций как в Y-хромосомах отдельных людей, так и их групп, племен, родов, популяций. Естественно, в данном контексте термин «популяций» относится к мужской половине рассматриваемой популяции. Методология новой науки — перевод динамической картины мутаций в хронологические показатели, во времена жизни общих предков популяций, а на самом деле — общих предков древних родов и племен. То есть фактически производится расчет времен, когда в древности жили эти рода и племена, и как эти времена меняются от территории к территории (Klyosov, 2009a, 2009b, 2009c; Klyosov, 2011; Rozhanskii, Klyosov, 2011; Klyosov, Rozhanskii, 2012; Klyosov, 2012).
Введем некоторые важные понятия и определения. Ниже они будут пояснены более детально, но важно их сразу обозначить. Во-первых, понятие «ДНК-генеалогия». Это – не генетика, в чем часто путаются даже профессионалы в науке. Генетика человека – это наука, изучающая наследственность и изменчивость признаков, определяющих врожденные особенности человека, и передаваемых, как правило, через гены. Но ДНК только на 2% состоит из генов, а в Y-хромосоме генов вообще ничтожная доля. Но дело даже не столько в этом. Направления и области науки определяются не объектами исследования, а методологией исследования. Именно методология исследования отличает, например, химию от физики, хотя объекты часто одинаковы. Методология генетики совершенно другая, чем методология ДНК-генеалогии. У них разные задачи исследований. ДНК-генеалогия в своей основе физическая химия, кинетика химических и биологических процессов, по сути задач – историческая наука, она оперирует хронологией, датировками древних событий, и для этого ДНК-генеалогия использует свой расчетный аппарат, которого нет в методологии генетики. ДНК-генеалогия, повторяю, использует и развивает методы физико-химической кинетики в применении к ДНК, переводит картину мутаций в хронологические, исторические показатели. Это вообще не входит в методологию генетики, у той другой экспериментальный и аппаратурный базис.
Ошибка – полагать, что направление науки определяется объектом исследования. Изучать ДНК – это далеко не обязательно генетика. Например, химик, растворяя ДНК в кислоте и изучая, скажем, вязкость получаемого раствора, вовсе не занимается генетикой. Альберт Сент-Дьорди как-то сказал – «Дайте химику динамомашину, и он тут же растворит ее в соляной кислоте». Это вовсе не означает, что химик при этом будет заниматься электродинамикой.
Некоторые, не слишком знакомые с предметом, полагают, что ДНК-генеалогия – это популяционная генетика. Это – совершенно не так. Популяционная генетика – это часть генетики, о чем говорит ее название, но со своими особенностями. Главная задача популяционной генетики – это выявление связи между генотипом и фенотипом, то, к чему ДНК-генеалогия не имеет отношения. Популяционная генетика зачастую тоже рассматривает гаплогруппы и гаплотипы Y-хромосомы, как и ДНК-генеалогия, но на этом сходство заканчивается. Аппарат популяционной генетики, применяемый для интерпретации получаемых данных, например, «метод главных компонент», характеризуется со стороны ДНК-генеалогии как совершенно примитивный и часто ведущий к заблуждениям, и не используется в ДНК-генеалогии. Он не отвечает задачам ДНК-генеалогии.
Это же относится и к искаженным и примитивным методам обработки картин мутаций в ДНК в рамках популяционной генетики. Если популяционный генетик увидит (или получит) набор из сотни гаплотипов, скажем, 37-маркерных, в котором, например, четыре гаплотипа одинаковых, и еще семь будут совпадать друг с другом случайными парами, он сообщит, что в наборе имеется «89 уникальных гаплотипов», и это и будет результатом его исследования, который пойдет в статью в научный журнал. Ни он, ни рецензенты не хотят признать, это эта «информация» не имеет ни малейшей ценности, и фактически никому в таком виде не нужна. Но таковы принципы и правила популяционной генетики. «Молекулярной историей» это назвать никак нельзя. А специалист в ДНК-генеалогии сразу скажет, что общий предок этих ста гаплотипов жил 925±105 лет назад, потому что натуральный логарифм отношения 100/4, поделенный на константу скорости мутации для 37-маркерных гаплотипов (0.09 мутаций на гаплотип на условное поколение в 25 лет) равен 36 → 37 условных поколений (стрелка здесь – поправка на возвратные мутации), то есть общий предок этой сотни гаплотипов жил примерно 925 лет назад. Сказать это с большей определенностью можно тогда, когда подсчитано число мутаций во всех ста гаплотипах, и если оно будет равно 324, тогда 324/100/0.09 = 36 → 37 условных поколений, то есть ровно тот же промежуток времени до общего предка. Погрешность расчетов определяется по известным правилам (Klyosov, J. Gen. Geneal., 2009a).
Совпадение времен до общего предка для «логарифмического» и «линейного» метода (в первом мутации не считаются, во втором считаются) свидетельствует, что закономерности образования мутаций в гаплотипах следуют кинетике первого порядка, что в свою очередь означает, что все сто гаплотипов действительно произошли от одного прямого общего предка.
Покажем это на примере, приведенном выше, для серии из 67 гаплотипов гаплогруппы R1a из Германии (см. рис. 1). В 12-маркерном формате из этой серии всего три гаплотипа идентичны друг другу, значит, это и есть базовые гаплотипы (гаплотипы 009, 011 и 013 на рис. 1). Получаем: [ln(67/3)]/0.02 = 155 → 183 условных поколений, то есть примерно 4575 лет до общего предка всей серии. Расчет по мутациям, которых в этой серии было 208 (см. выше), дает 208/67/0.02 = 155 → 183 условных поколений (по 25 лет) от общего предка, то есть 4575±560 лет до общего предка. Совпадение получилось практически абсолютным, но это в данном расчете случайно, потому что чисто статистически в данной серии могли оказаться как 2, так и 4 базовых гаплотипа, и погрешность при таких малых количествах базовых гаплотипов велика. Тем не менее, логарифмический (без подсчета мутаций) и линейный (с подсчетом мутаций) методы расчетов показывают, что серия гаплотипов вполне подходит для расчетов, и что общий их предок действительно жил примерно 4600 лет назад.
Но для 25-маркерных гаплотипов при таком малом числе гаплотипов и таком довольно удаленном общем предке расчет по логарифмическому методу уже невозможен. Даже два базовых 25-маркерных гаплотипа сохраняются всего лишь на протяжении [ln(67/2)]/0.046 = 76 → 83 условных поколений, то есть примерно 2075 лет. После этого и двух базовых гаплотипов не остается. Но даже отсюда можно – просто на взгляд, без расчетов – получить граничную информацию о времени жизни общего предка серии гаплотипов, в данном случае – больше 2100 лет назад. Здесь 0.02 и 0.046 – константы скорости мутаций для 12- и 25-маркерных гаплотипов.
Популяционная генетика такими подходами не пользуется, они ей чужды по ряду причин, среди них – приверженность устаревшим, традиционным подходам, отсутствие соответствующего образования, клановость, отсутствие хорошей научной школы. Короче, ДНК-генеалогия является другой дисциплиной, нежели популяционная генетика.
Рассмотрим понятие «молекулярная история». Суть его в том, что становится возможным следить за передвижениями древних народов не с помощью лопаты и кисточки археолога, не обмеряя черепа, не хитроумно расплетая созвучия и значения слов в живых и мертвых языках, не изучая древние фолианты в библиотеках и монастырях, а просто прослеживая за метками в Y-хромосомах в наших ДНК. Они, эти метки, не могут «ассимилироваться», или «поглотиться» другими языками, культурами, или народами, как это происходит тысячелетиями в рамках понятий истории, лингвистики, антропологии. Иначе говоря, методология новой исторической науки основывается на изучении молекул нуклеиновых кислот, а именно ДНК, дезоксирибонуклеиновой кислоты. То, что еще несколько лет назад казалось каким-то развлечением, оказалось, дает истории, антропологии, археологии, лингвистике возможность проверить концепцию, рассмотреть данные под принципиально другим углом, связать воедино казалось бы разрозненные части общей картины наших знаний об окружающем мире. Разумеется, не следует понимать буквально слова выше, как то, что отныне не нужно читать фолианты, изучать языки и не надо проводить археологических раскопок. Напротив, это все приобретает еще бóльшую значимость и ценность, поскольку усиливает выводы традиционных подходов в совокупности с выводами ДНК-генеалогии, при условии взаимного их согласования. Или, напротив, заставляет пересматривать казалось бы устоявшиеся выводы, если они входят в явное противоречие с данными ДНК-генеалогии.
Дадим еще пример принципиального различия ДНК-генеалогии и популяционной генетики. Ниже мы покажем на отдельном примере анализ бактрийских гаплотипов, предположительно предковых к гаплотипам современных карачаево-балкарцев и осетин. Мы постоили дерево гаплотипов, разделили его на ветви (точнее, сам вид дерева уже показал четкое разделение на ветви), определили базовый (то есть предполагаемый предковый) гаплотип для каждой ветви, рассчитали времена для общих предков каждой ветви, провели явные параллели с осетинами и карачаево-балкарцами, провели перекрестные расчеты и показали, что они согласуются со временами жизни общих предков бактрийских гаплотипов, как и современных пуштунов Афганистана и Пакистана. Это в свою очередь позволило предложить гипотезу в отношении картины и времен миграций носителей субклада G2a1 из Бактрии на Кавказ.
Посмотрим, что, имея такие же исходные данные, получают из них популяционные генетики. Эти исходные данные (гаплотипы) были опубликованы в статье (Lee et al, 2014), и, поскольку статья совсем недавняя, отражает современные подходы популяционной генетики. Какие же главные выводы сделали авторы из опубликованного ими материала? Помогает то, что авторы приложили к статье список highlights, то есть главных выводов. Их – пять: (1) определили гаплотипы для 270 человек, (2) «дискриминационная способность» для 230 гаплотипов из всех 270 была равна 85.2%, (3) три маркера, DYS446, DYS447 and DYS449, были основными в отношении «увеличения дискриминации», (4) доминирующая гаплогруппа среди пакистанских пуштунов была R1a1a, (5) предками пакистанских пуштунов вряд ли были евреи. Резюме к статье добавляет, что (1) каждый маркер показывает «разнообразие» от 0.2506 до 0.8538, (2) «дискриминационная способность» для 17-маркерных гаплотипов была равна 73.7%, и при увеличении их длины возросла до 85.2%, (3) для получения информации о происхождении пуштунов сравнили носителей гаплогрупп G и Q у пуштунов и евреев (ашкенази и сефардов), но еврейского происхождения пуштунов особенно не обнаружили («little support for a Jewish origin could be found»). Все эти «разнообразия маркеров» считаются без разделения соответствующих гаплотипов на гаплогруппы, и на круг по выбранным 199 гаплотипам «разнообразие» получилось равно 0.9903, а по 230 гаплотипам оно было равно 0.9968. Ни для чего больше эти цифры в статье не использовались, датировки не считались. Да и не могли считаться, потому что для них все гаплогруппы «сваливались в кучу». Никакого практического, и тем более теоретического смысла все эти арифметические упражнения популяционных генетиков не имели и иметь не могут. Смысл там исключительно описательный, что увидели, то и записали.
В этих примерах – суть подходов и уровня работы популяционных генетиков. Теперь оцените высказывание проф. Е. Балановской с трибуны недавней Конференции, что «Клёсов эксплуатирует собранную генетиками информацию, обрабатывает её неизвестными методами и получает красивые результаты на потребу публике». Так если попгенетики беспомощны в корректной и информативной обработке собранных ими гаплотипов, то надо заменить их некомпетентность и неумение на полученные корректными методами «красивые результаты», тут Балановская не ошиблась. А что до «неизвестных методов», то учиться надо. И придется учиться, деваться просто некуда. А уж «на потребу публике» – это ложь Балановской от беспомощности и отчаяния, от своей некомпетентности. Красивые результаты – это правильные результаты, что еще физики давно подметили.
Различия терминов в ДНК-генеалогии и популяционной генетике
Поскольку ДНК-генеалогия и популяционная генетика – разные области науки, с разными методологиями, разным характером выводов, разными целями и задачами, разным способом подачи данных, то у них, разумеется, во многом разная терминология. От страстного желания хоть как-то дискредитировать, Балановская объявила с трибуны Конференции, что я путаю термины. Да не путаю я, я просто использую другие термины, термины ДНК-генеалогии. Она так и не поняла, что ДНК-генеалогия – другая наука, нежели попгенетика с ее терминами. Например, предковый гаплотип в ДНК-генеалогии называется «базовым», потому что он вычисляется, и нет гарантии, что он действительно предковый. Возможно, отклоняется на одну аллель. Зачем вводить читателей в заблуждение?
В ДНК-генеалогии используются поправки на возвратные мутации, которых нет в популяционной генетике. В ДНК-генеалогии есть логарифмический метод, кратко описанный ранее. И он вовсе не «неточный», как безграмотно заявил О. Балановский, в очередной раз показав свою некомпетентность. Неточный этот метод бывает тогда, когда используется неправильно. Неточным будет микроскоп, если его использовать для рассматривания звезд. На то есть телескоп, или хотя бы подзорная труба.
Чем больше по численности серия гаплотипов, происходящих от одного предка, тем расчет времени жизни этого предка точнее. И тем больше экономия времени при расчетах, поскольку логарифмический метод не требует подсчета числа мутаций в гаплотиипах. Например, из 2000 12-маркерных гаплотипов субклада R1b-L21 114 гаплотипов являются базовыми, то есть идентичными друг другу. Это дает [ln(2000/114)]/0.02 = 143 → 167 условных поколений, или 4175±570 лет до общего предка. Расчет по мутациям дает 4325±350 лет до общего предка. Разница между логарифмическим и линейным (то есть по мутациям) методами расчета дает всего лишь 3.6% разницы, хотя погрешность расчетов ее увеличивает. Если округлить полученные величины с учетом погрешностей до 4200 и 4300 лет до общего предка, разница между ними становится равной всего лишь 2.4%.
Ну, и где здесь «неточный» логарифмический метод, при разнице 2-3% с линейным? Язык у Балановского без костей, как говорят в народе. И это говорит тот, ошибки у которого в расчетах датировок систематически равны как минимум 250-350%.
В ДНК-генеалогии используются понятия констант скоростей мутаций, а не просто «скоростей», как в попгенетике. Последнее в данном контексте неверно.
В ДНК-генеалогии нет «эффекта основателя», потому что этот термин излишний, он не нужен при правильной обработке данных. В ДНК-генеалогии арии – это древние носители гаплогруппы R1a, и наоборот, древние носители гаплогруппы R1a – это арии. Хотя бы потому, что носители гаплогруппы R1a составляют до 72% высших каст Индии (Sharma et al., J. Human Gen., 2009). И еще потому, что слово арии созвучно индексу R1a. По той же причине древние носители гаплогруппы R1b – это эрбины, поскольку их называть как-то надо, а название отсутствует. Эрбины – это хороший термин, простой и ясный. Поэтому, читая фразу «Балановский особо подчеркнул вызывающий недоумение случай создания в рамках «ДНК-генеалогии» фантомного этноса «эрбинов», якобы представляющих собой гаплогруппу R1b», опять приходится сомневаться в способности критика соображать. О терминах, как известно, не дискутируют, их или принимают, или не принимают. Недоумение при этом выказывают только люди недалекие, что и имеем. Далее, эрбины – это вовсе не «этнос», похоже, что популяционный генетик Балановский не знает, что гаплогруппа этнос не определяет. И эрбины не «якобы» представляют гаплогруппу R1b, а буквально, по определению.
Наконец, в терминах ДНК-генеалогии гаплогруппа – это род. В основе как гаплогруппы, так и рода (это синонимы) находился патриарх, родоначальник, потомки которого выжили cо своими гаплотипами до настоящего времени.
Итак, вывод: в ДНК-генеалогии своя терминология, отличная от таковой в популяционной генетике.
Расчётный аппарат ДНК-генеалогии. Калибровка. Примеры.
Читаем «мою» страничку в Википедии. Там недавно появилось следующее: «Ряд известных российских антропологов и генетиков выступил с критикой Клёсова и развиваемой им «ДНК-генеалогии». Так, Олег Балановский, заведующий лабораторией геномной географии Института общей генетики РАН, полагает, что технические методы «ДНК-генеалогии» вовсе не оригинальны, представляя собой обычный метод молекулярных часов с рядом простейших модификаций. …С его точки зрения, технические аспекты «ДНК-генеалогии» являются «малоэффективными, но приемлемыми»…».
Сказана глупость, и не одна, а несколько всего в нескольких строках.
Во-первых, что такое «обычный метод молекулярных часов»? Это – шаблонная фраза, за которой нет никакой мысли, обычное попугайское повторение того, что когда-то услышал. «Метод Животовского» – тоже «обычный метод молекулярных часов», только часы торопятся в три раза, давая датировки втрое завышенные. Любая реакция в химических или биологических системах, описываемая константой скорости первого порядка, есть «обычные молекулярные часы», поскольку связана с хронологией процесса на молекулярном уровне. Динамика любого такого процесса связана со временем согласно формуле с = с0е-kt, где с0 – исходное состояние системы (например, начальное количество или концентрация изучаемого вещества; количество гаплотипов Y-хромосомы в изучаемой выборке, и т.д.),с – состояние системы в определенный момент времени t (где t – время прошедшее с начала реакции, t-t0), или количество базовых, то есть исходных гаплотипов в изучаемой выборке в настоящее время, спустя время t, прошедшее со времени жизни общего предка изучаемой серии гаплотипов), k – константа скорости реакции. Эту же формулу можно переписать в виде ln(с0/c) = kt, и она становится выражением логарифмического метода анализа выборок гаплотипов в ДНК-генеалогии. Берем, скажем, сто или тысячу гаплотипов, или любое другое их число, делим на число базовых (то есть одинаковых, идентичных друг другу гаплотипов, суть предковых гаплотипов, которые не успели мутировать за время t, прошеднее со времени жизни общего предка), берем натуральный логарифм (ln), и получаем произведение kt, то есть константу скорости мутации, помноженную на число лет, прошедшее со времени жизни общего предка, или на число условных поколений, опять же прошедших после общего предка – в зависимости от того, выражали константу скорости в годах, или в поколениях.
Отсюда уже видно, что неважно, сколько лет положить на условное поколение – 20, 25, 30, 35 или любое другое число лет, поскольку константа скорости мутации тут же подстроится, они же завязаны друг на друга, произведение-то одно. В ДНК-генеалогии берется 25 лет на условное поколение, и, соответственно, константы скорости мутации приобретают определенные значения, получаемые по калибровке (см. ниже). Например, для 12-маркерных гаплотипов константа равна 0.02 мутаций на гаплотип на условное поколение, для 25-маркерных 0.046 мутаций на гаплотип на условное поколение, для 37-маркерных – 0.09, для 67-маркерных – 0.12, для 111-маркерных – 0.198 мутаций на гаплотип на условное поколение. Если это пересчитать в расчете не на гаплотип, а на маркер, то получим соответствующие константы скорости 0.00167, 0.00184, 0.00243, 0.00179, 0.00178 мутаций на маркер на условное поколение. Уже видно, что константы скорости разные для разных гаплотипов, и различаются, например, для 37-маркерных и 12-маркерных гаплотипов в 1.46 раз, то есть на 46%. А если сравнить с 6-маркерными гаплотипами (константа скорости мутации на гаплотип равна 0.0088, на маркер 0.00147), то диапазон различий в константах в зависимости от длины маркера расходится уже на 1.65 или на 65%. Вывод – никак нельзя принимать константы скорости мутации на маркер за постоянные величины, одинаковые для всех гаплотипов. 65% ошибки в расчетах – цена такого неумного (или неквалифицированного, или некомпетентного) предположения.
А что делают популяционные генетики? Именно так и принимают. Что константы скорости мутации (они говорят «скоростей мутации», без «констант», что технически неверно) для всех гаплотипов одинаковы. Более того, что они – «по Животовскому» – равны 0.00069 на маркер на 25 лет, для всех гаплотипов без разбору. Ошибку такого неквалифицированного подхода легко посчитать – она гуляет от гаплотипа к гаплотипу другой протяженности в диапазоне от 213% ошибки до 352%, и это только в «чистом случае», когда выборка гаплотипов симметрична. Если несимметрична – можно легко уйти в значительно большие ошибки. Так работает попгенетика. Это и есть псевдонаучный подход. Хотя – тоже «молекулярные часы», только никуда не годятся, время неправильно показывают.
С «молекулярными часами» разобрались, доктор наук Балановский не имеет об этом никакого понятия. Он не понимает, что иметь часы – дело нехитрое, но надо, чтобы они были отрегулярованы. Это означает, что к ним должны прилагаться корректные константы скоростей мутаций. С этим у попгенетиков просто драма, которая тянется уже десятилетия, а с появлением «скоростей Животовского», они же «популяционные скорости», в 2004 году, драма стала трагедией. Вот где пошла псевдонаука. Об этом – в следующем разделе.
Но перед этим («во-вторых», продолжая «во-первых» выше) – другие глупости, которые от вопиющей малограмотности объявил новоиспеченный доктор наук Балановский. Впрочем, мы о них уже упомянули. Он объявил, что «технические методы «ДНК-генеалогии» вовсе не оригинальны, представляя собой обычный метод молекулярных часов с рядом простейших модификаций. …С его точки зрения, технические аспекты «ДНК-генеалогии» являются «малоэффективными, но приемлемыми»…».
С «молекулярными часами», повторяю, разобрались. На что ни пойдет безграмотный человек, чтобы дискредитировать грамотного… пустится во все тяжкие, включая и позор на публике. Так что же такое «ряд простейших модификаций»? Введение впервые в оборот серии констант скоростей мутаций, нигде и никогда раньше не публиковавшихся? Введение в оборот логарифмического метода, совершенно незнакомого ранее попгенетикам? Введение в оборот поправок на возвратные мутации, о которых попгенетики говорили, но никто и никогда не рассчитывал и не применял? Введение в оборот калибровок констант скоростей мутаций (об этом чуть ниже)?
Не знаю как для кого, а для меня ясно, что О. Балановский, будучи «на автопилоте» желания дискредитировать, себя уже не контролировал. Он что-то произносил, главное, чтобы было негативное, но смысла сам не понимал. Это – не наука, это и есть лженаука в исполнении О. Балановского, как и другого доктора, Е. Балановской.
Самое забавное в том, что этот незнайка, применяя «методы» популяционной генетики и ошибаясь на сотни процентов в датировках, заявляет, что «технические аспекты «ДНК-генеалогии» являются «малоэффективными, но приемлемыми»…». «Малоэффективные» произнесено на том же автопилоте, в том же раже дискредитировать. Если бы его спросили расшифровать понятие «малоэффективные», он бы не нашел, что ответить. Ни одного примера привести бы не смог. За «приемлемые» – отдельное спасибо, мой сарказм заметен?
Переходим к калибровке, а именно, как были получены те самые константы скоростей мутаций, о которых попгенетика не имела понятия, да и сейчас не имеет. Литературу они или не читают, или читают от сих до сих. Посмотрим еще раз на «мою» страничку в Википедии. Там – следующее: «Елена Балановская, заведующая лабораторией популяционной генетики человека Медико-генетического научного центра РАМН, отмечает, что Клёсов эксплуатирует собранную генетиками информацию, обрабатывает её неизвестными методами». Мало того, что я в основном использую общественные базы данных по гаплотипам, предоставляемых коммерческими компаниями, или это тоже «эксплуатация информации»? Оборот-то какой, уже с идеологическим уклоном. Правда, как злонамеренный, так и бессмысленный, информацию нельзя «эксплуатировать». Но до зловонной кучи, оставленной Балановскими, все сгодится. Меня, кстати, засыпают письмами, почему я не удалю сам эту чепуху со странички в Википедии? На это я отвечаю, что не я эту страничку создавал, не мне ее и менять. Там много погрешностей, вплоть до неверного места моего рождения. Пусть как создали, так и будет, я памятники себе не создаю и любовно не подправляю. А в отношении мусора, который туда вбросили Балановские и их подельники, так пусть их дурь каждому видна будет.
Но я сейчас – про «обрабатывает неизвестными методами». Занятно, не так ли? Эти методы публикуются с 2008 года, они опубликованы на 14 страницах журнала Human Genetics еще в 2009 году, они опубликованы в старейшем академическом журнале «Биохимия» (Москва), в дюжине других научных статей, в основном англоязычных, в книгах, они активно обсуждались на международных и российских сайтах – на один, сайт «Родство», кстати, захаживал Балановский, мог бы прочитать и маме рассказать. Но он предпочел там устроить скандал, называя гаплогруппу R1a фашистской, потому что она «арийская», за что и был выставлен с сайта модератором П. Шваревым. Сейчас Балановский рассказывает с трибуны Конференции по карачаево-балкарцам трогательную историю, что его выставили с сайта только потому, что у него другая гаплогруппа, не как у остальных. Лжец он и есть во всем.
Так вот, про калибровки. Эти данные были опубликованы еще в 2011 году (Rozhanskii & Klyosov, Advances in Anthropology, vol. 1, No. 2, 26-34), и недавно изложены И.Л. Рожанским на Переформате. Так что буду краток, желающие могут прочитать подробности на английском или русском языках. Суть в том, что были взяты генеалогические данные для 13 семей, удовлетворяющие сформулированным жестким критериям. Эти 13 семей (или «Проектов») были отобраны из сотен других, которые были менее многочисленны или датировки которых были менее достоверны, или гаплотипы были короткими, то есть низкого разрешения.
В указанной статье в Advances in Anthropology приведены многочисленные графики для гаплотипов разной протяженности, и каждый график иллюстрировал надёжность калибровки, доверительные интервалы и прочее. Статья – редкая по глубине обоснований и достоверности полученных данных, результаты калибровок выверены на 3160 гаплотипах из 55 гаплогрупп и субкладов, из них 2489 гаплотипов были 67-маркерными, с которыми попгенетики не только не работают, но не могут и мечтать. После этого смешно читать высказывания Балановских – на том же негативном автопилоте – что «журнал не индексирован, он с претензией на научность». Воистине, злоба и злонамеренность границ не имеют. Опять, вопиющее нарушение научной этики Балановскими. В науке не индексирование обсуждают, а качество, надежность данных, глубину и обоснованность выводов. Мне, как главному редактору журнала (хотя в том 2011 году я главным редактором не был), не индексированность нужна, а то, что журнал читают десятки тысяч людей, как следует из раздела «наиболее популярные статьи журнала» (ссылка). А лаборантам от науки, которыми Балановские являются, нужны «шашечки», хотя сами в «индексированных» журналах производят обильный мусор.
Не буду приводить все калибровочные графики, дам только калибровочную диаграмму для 37- и 67-маркерных гаплотипов. Здесь на горизонтальной оси – число лет до общего предка каждой из ДНК-генеалогических «семей», на вертикальной оси – среднее число мутаций на маркер (поскольку гаплотипы разные – 37- и 67-маркерные) в гаплотипах, принадлежащих этим семьям.
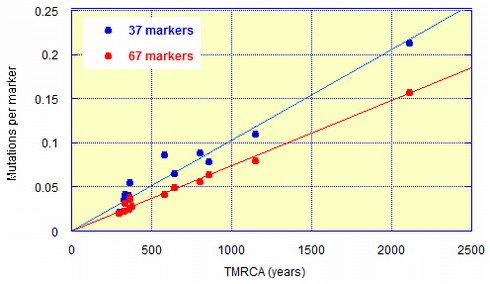
Рис. 3. Калибровочные взаимоотношения между временем, прошедшим от общих предков генеалогических серий гаплотипов в каждой группе («генеалогической семье»), и числом мутаций, накопившихся с того времени, в расчете на маркер в гаплотипах в 37- и 67-маркерном формате. Данные и принцип подхода опубликованы в журнале Advances in Anthropology (2011, Rozhanskii & Klyosov) и приведены в дополненном виде И.Л. Рожанским на Переформате.
Видно, что есть четкая линейная зависимость между «возрастом» ДНК-линии и числом мутаций на маркер. Так и должно быть, поскольку мутации в гаплотипах (и маркерах) происходят случайным образом и описываются кинетикой первого порядка. Угол наклона корреляционной прямой для 37-маркерных гаплотипов выше, чем у 67-маркерных, поскольку мутации в 37 маркерах (где большая доля «быстрых» по мутациям маркеров) в совокупности происходят чаще, чем в 67-маркерных (где, напротив, маркеры с 38 до 67-го по порядку содержат высокую долю «медленных» маркеров). Отсюда и получились те константы скоростей мутаций, приведенные выше в данной статье: для 37-маркерных гаплотипов 0.00243 мутаций на маркер (0.09 мутаций на гаплотип) за условное поколение, для 67-маркерных гаплотипов 0.00179 мутаций на маркер (0.12 мутаций на гаплотип) за условное поколение. Для 67-маркерных гаплотипов точность калибровки больше, и соответствует погрешности в определении константы скорости мутаций ±2.5 % (И.Л. Рожанский, ссылка приведена выше). Рассмотрение тех нескольких тысяч гаплотипов, упомянутых выше, из 55 гаплогрупп и субкладов (Advances in Anthropology, 2011) позволило заключить, что использованная нами калибровка и метод расчета дают точность в определении времени жизни предка с точностью ±10 % или меньше в интервале от 500 до 6000 лет назад.
Для сравнения – у популяционных генетиков датировки получаются с ошибками 250-350% (и выше), да и там с погрешностями ±50-80%. Вот это и есть псевдонаука, она же лже-наука. Псевдо- и лже- не потому что ошибаются, ошибки в науке нормальное дело, наука так развивается, а потому что уже по меньшей мере пять лет нормальные, корректные, обоснованные подходы ДНК-генеалогии уже опубликованы, в упомянутой статье 2011 года они просто более основательно уточнены (а фактически подтверждены), но популяционная генетика упрямо эти данные игнорирует, и продолжает идти по лже-пути, генерируя мусор в академических журналах, повторю это еще не один раз. Вот почему попгенетика в отношении датировок, что является, видимо, важной частью их «исследований», и соответствующих исторических «интерпретаций», является псевдо-наукой. А поскольку «псевдонауки» не бывает, как и осетрины второй свежести, то это просто ложь и обман. На том Балановские и иже с ними и живут, ложь и обман – их и питательная почва, и хлеб. Это же в полной мере относится к пасквилям, которые они в последнее время активно вбрасывают в СМИ.
Дам еще иллюстративный пример. В Шотландии есть знаменитая «генеалогическая» семья МакДоналдов, их несколько тысяч человек, на них работает целый штат профессиональных генеалогов, их документальная генеалогия изучена во всех возможных деталях. И неудивительно, они берут свое начало от шотландского вождя Сомерледа, среди их предков наполеоновский маршал МакДоналд и много других выдающихся людей. Основная группа МакДоналдов гаплогруппы R1a ведет свою линию от Джона Лорда Островов (John Lord of the Isles), умершего в 1386 году, то есть 628 лет назад. Принимая 25 лет на условное поколение (именно эта величина входит в величины констант скоростей мутаций, приведенные выше), получаем, что Джон умер 25 условных поколений назад, то есть жил примерно 26 условных поколений назад. В своей книге «Происхождение человека» (2010) я рассмотрел серию гаплотипов потомков Джона, которая тогда насчитывала сначала (в 2008 году) 68 гаплотипов, затем (в 2010 году) 84 гаплотипа. Среди 68 гаплотипов в 6-маркерном виде было 17 мутаций, и 53 гаплотипа были немутированными, то есть базовыми. Считаем по правилам ДНК-генеалогии. Сначала по мутациям, используя так называемый линейный метод: 17/68/0.0088 = 28±7 условных поколений до Джона (погрешность велика, поскольку мутаций мало). Теперь считаем без учета мутаций, используя логарифмический метод: [ln(68/53)]/0.0088 = 28±5 условных поколений назад. 0.0088 – это константа скорости мутаций для 6-маркерных гаплотипов.
Потом появились 60 25-маркерных гаплотипов, в них – 69 мутаций, и из них 18 гаплотипов были базовыми, то есть идентичными друг другу. Получаем:
Линейный метод – 69/60/0.046 = 25±4 условных поколений.
Логарифмический метод – [ln(60/18)]/0.046 = 26±7 условных поколений.
Потом, с расширением тестирования, появились 84 25-маркерных гаплотипов, и в них во всех содержалось 109 мутаций. В тех же 84 гаплотипах в 12-маркерном формате было 44 мутации. Получаем:
Для 12-маркерных гаплотипов 44/84/0.02 = 26±5 условных поколений.
Для 25-маркерных гаплотипов 109/84/0.046 = 28±4 условных поколений.
Здесь 0.02 и 0.046 – это константы скорости мутации для 12- и 25-маркерных гаплотипов, соответственно. Итак, наблюдается совершенно разумное совпадение с документальной генеалогией по Джону и его потомкам: документальная генеалогия дает 628 лет до годовщины смерти Джона (1386 год), а ДНК-генеалогия дает 650-700 лет до времени его жизни (1314-1364 год), то есть практически абсолютное совпадение в контексте данного исследования, учитывая, что год его рождения неизвестен.
И таких примеров десятки, если не сотни, когда ДНК-генеалогия дает либо совпадения с документальной генеалогией (в пределах погрешности расчетов), либо дополняет документальную генеалогию, когда ее данные фрагментарные или вовсе отсутствуют, кроме приблизительных датировок.
Продолжение следует…
Анатолий А. Клёсов,
доктор химических наук, профессор
")
")
")